
A foggy day and cold as hell. Got 8 flights delayed and 1 flight cancelled.




 The female of the dreaded gypsy moth
The female of the dreaded gypsy moth Diptera
Diptera Halteres on a cranefly
Halteres on a cranefly Worker ants have lost their wings, but not the capacity to grow wings. They lost their wings in evolution because they are a nuisance and get in the way underground. Queen ants and male ants have wings and worker ants are females who could have become queens but for environmental reasons (not genetic) failed to become queens. Queen ants use their wings only once, to fly out of the natal nest, find a mate and then settle down to dig a hole for a new nest. As they begin their new life underground the first thing they do is lose their wings, in some cases by literally biting them off.
Worker ants have lost their wings, but not the capacity to grow wings. They lost their wings in evolution because they are a nuisance and get in the way underground. Queen ants and male ants have wings and worker ants are females who could have become queens but for environmental reasons (not genetic) failed to become queens. Queen ants use their wings only once, to fly out of the natal nest, find a mate and then settle down to dig a hole for a new nest. As they begin their new life underground the first thing they do is lose their wings, in some cases by literally biting them off. Ostrich Wing Stubs are only used for balancing and steering while running and when they enter into social and sexual displays.
Ostrich Wing Stubs are only used for balancing and steering while running and when they enter into social and sexual displays. Kiwi wings are too small to be seen outside the birds fine coat of feathers, but vestiges of wing bones are there.
Kiwi wings are too small to be seen outside the birds fine coat of feathers, but vestiges of wing bones are there.  Moas (which got extinct 1500AD)
Moas (which got extinct 1500AD) Kakapos, the heavyweight champion of all parrots
Kakapos, the heavyweight champion of all parrots  Galapagos Cormorants
Galapagos Cormorants blowhole of a dolphin
blowhole of a dolphin  The skeleton of a whale. Notice (c) the hind legs that have no utility otherwise
The skeleton of a whale. Notice (c) the hind legs that have no utility otherwise whale brain
whale brain human brain
human brain fish brain
fish brain Dungongs
Dungongs whales
whales Instead of spinning regular silk webs, they form platicy, silky air bubbles on the surface of water
Instead of spinning regular silk webs, they form platicy, silky air bubbles on the surface of water  2. Alignment (steer to the average heading of local flockmates)
2. Alignment (steer to the average heading of local flockmates)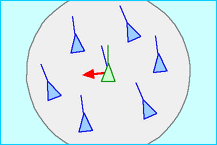 3. Cohesion (steer to move towards the average position of local flockmates)
3. Cohesion (steer to move towards the average position of local flockmates) Each boid has direct access to the whole scene's geometric description, but flocking requires that it only interacts with flockmates within a certain small neighbourhood around itself. The neighbourhood is characterized by distance (measured from center of the boid) and an angled (measured from the direction of boid's flight). Flockmates outside this neighbourhood are ignored.
Each boid has direct access to the whole scene's geometric description, but flocking requires that it only interacts with flockmates within a certain small neighbourhood around itself. The neighbourhood is characterized by distance (measured from center of the boid) and an angled (measured from the direction of boid's flight). Flockmates outside this neighbourhood are ignored. The key point is that there is no choreographer and no leader. Order, organization, structure - these all emerge as by-products of rules which are obeyed locally and many times over, not globally. And that is how embryology works. It is all done by local rules, at various levels but especially the level of the single cell. In the field of development, or manufacture, the equivalent of this kind of programming is self-assembly.
The key point is that there is no choreographer and no leader. Order, organization, structure - these all emerge as by-products of rules which are obeyed locally and many times over, not globally. And that is how embryology works. It is all done by local rules, at various levels but especially the level of the single cell. In the field of development, or manufacture, the equivalent of this kind of programming is self-assembly. Domesticated Silver Fox
Domesticated Silver Fox




The human-faced crabs in Japan from Fauna Japonica. Crustacea. (detail of the Heike-gani in the right)
From the top: Heike-gani, Samehada-heike-gani (Shark-skin Taira-clan’s crab), Kimen-gani (Devil’s mask crab)
Kyoto University Library in Japan
The species of the crab known as Heikea Japonica is found in Japanese waters. The generic name, Heikea, comes from a japanese clan called the Heike, who were defeated at sea in the battle of Danno-Ura (1185) by a river clan called Genji. Legend tells that the ghosts of drowned Heike warriors now inhabit the bottom of the sea, in the bodies of crabs - Heikea Japonica. The myth is encouraged by the pattern on the back of the this crab, which resembles the fiercely grimacing face of a samurai warrior










{kind=link}
{kind=link}